Ionizing radiation consists of high-energy particles such as
photons or ions,
whose tracks deposit energy in a cell [20, 21].
Ionizing radiation
dose, ![]() , is measured in energy per unit mass, with 1 Gy
being 1 Joule/kg. The time dependence of dose
is much more precisely defined than in chemotherapy,
as pharmacokinetic or other accessibility issues do not really
arise. A complication in some cases is the stochastic nature
of radiation (survey in [20]). However most clinical
applications
involve sparsely ionizing radiations (such as
, is measured in energy per unit mass, with 1 Gy
being 1 Joule/kg. The time dependence of dose
is much more precisely defined than in chemotherapy,
as pharmacokinetic or other accessibility issues do not really
arise. A complication in some cases is the stochastic nature
of radiation (survey in [20]). However most clinical
applications
involve sparsely ionizing radiations (such as ![]() -rays
or hard x-rays) at doses above 1 Gy; in these cases
so many independent particles (e.g. photons) strike a cell
nucleus that dose, a deterministic quantity by definition,
suffices to
quantify the insult. We shall here always suppose irradiation
occurs within the finite time interval
-rays
or hard x-rays) at doses above 1 Gy; in these cases
so many independent particles (e.g. photons) strike a cell
nucleus that dose, a deterministic quantity by definition,
suffices to
quantify the insult. We shall here always suppose irradiation
occurs within the finite time interval ![]() .
.
![]() denotes accumulated dose at time
denotes accumulated dose at time ![]() .
Thus
.
Thus
![]() is radiation dose rate, here assumed
smooth.2
is radiation dose rate, here assumed
smooth.2
The most important radiation
damage is to chromatin, e.g. DNA double strand breaks (DSBs).
An acute dose of 1 Gy makes many thousands of ionizations
in the cell's nucleus, of which a small minority, ![]() 40
in a human cell, quickly induce DSBs.
Most DSBs are repaired during the next half hour or so,
and a few are misrepaired. Many of the misrepairs
involve a binary reaction between two different DSBs
[22].
At a typical dose, of several Gy,
at least one misrepair usually occurs, which can be
enough to kill the cell at the next mitosis.
Other kinds of clonogenically lethal lesions, formed "directly" by
irradiation and not subject to repair or misrepair (e.g. lethal
point mutations) can also be present.
40
in a human cell, quickly induce DSBs.
Most DSBs are repaired during the next half hour or so,
and a few are misrepaired. Many of the misrepairs
involve a binary reaction between two different DSBs
[22].
At a typical dose, of several Gy,
at least one misrepair usually occurs, which can be
enough to kill the cell at the next mitosis.
Other kinds of clonogenically lethal lesions, formed "directly" by
irradiation and not subject to repair or misrepair (e.g. lethal
point mutations) can also be present.
The LQ model
has been derived from many different points of view, and is
actually more robust than any one of its derivations. It has
been applied to cell killing, chromosome aberration formation,
mutation, transformation and other endpoints. For brevity we
here confine attention to cell killing
in the sense of clonogenic inactivation, and to tumor radiotherapy.
The applications to endpoints more directly related to
carcinogenesis, such as chromosome aberrations,
are, broadly speaking, similar.
One derivation of the model starts from a standard pair of ODEs
[23], as
follows. The dependent functions
are the average number ![]() of DSBs
per cell at time
of DSBs
per cell at time ![]() and the average number
and the average number ![]() of cells per
population (i.e. per tumor or cell culture). The ODEs are:
of cells per
population (i.e. per tumor or cell culture). The ODEs are:
It often happens, for doses less than about 5 Gy, that
one can approximate by setting
![]() in Eq. 8
(however, the related term involving
in Eq. 8
(however, the related term involving ![]() may be important
in Eq. 9
even when the binary,
may be important
in Eq. 9
even when the binary, ![]() , term is
negligible compared to the other terms in in Eq. 8).
If
, term is
negligible compared to the other terms in in Eq. 8).
If
![]() , Eq. 8 can
be explicitly integrated. For times greater than
the radiation stoppage time
, Eq. 8 can
be explicitly integrated. For times greater than
the radiation stoppage time
![]() and large compared to the repair time
and large compared to the repair time ![]() , i.e. for
, i.e. for
![]() , which we abbreviate by
, which we abbreviate by ![]() , inserting the integral
of Eq. 8 into 9 and integrating again gives
the LQ model. Specifically the result of the two integrations
is:
, inserting the integral
of Eq. 8 into 9 and integrating again gives
the LQ model. Specifically the result of the two integrations
is:
The most essential, virtually ubiquitous
features characteristic of ionizing radiation damage,
captured here in Eqs. 10 and 11,
are the following: part of the damage is dose-rate independent
and (approximately) linear in dose (term involving ![]() );
part of the damage corresponds to
misrepair of repairable damage (term involving
);
part of the damage corresponds to
misrepair of repairable damage (term involving ![]() );
this part gives rise to effects super-linear in dose
(here
);
this part gives rise to effects super-linear in dose
(here
![]() ); and this part is decreased by prolonging
the dose instead of giving dose all at once.
The fact that the LQ model quantifies these normally dominant
effects, and, equally important, neglects or slurs over a host of
less important effects so that only three adjustable
parameters are involved, accounts in large part for its widespread
recent popularity.
At worst, the model summarizes, in a highly condensed quantitative
form,
a truly enormous body of radiobiological data and clinical experience.
); and this part is decreased by prolonging
the dose instead of giving dose all at once.
The fact that the LQ model quantifies these normally dominant
effects, and, equally important, neglects or slurs over a host of
less important effects so that only three adjustable
parameters are involved, accounts in large part for its widespread
recent popularity.
At worst, the model summarizes, in a highly condensed quantitative
form,
a truly enormous body of radiobiological data and clinical experience.
The detailed properties of the repairable damage component have long been controversial, and a wide variety of ODE models other than Eqs. 8 and 9 have been proposed (survey in [24]). For example, many authors have suggested saturation of repair enzymes rather than binary misrepair as an underlying molecular mechanism [29]. Remarkably, however, first order time-dependent perturbation theory shows that the LQ model, Eqs. 10 and 11, is a low-dose/intermediate-dose approximation to virtually all the different ODE models [30]. There are presumably many different damage repair/misrepair pathways simultaneously operative in a cell, but the LQ model approximates most of them.
The model has become the tool of choice for quantifications of treatment planning in tumor radiotherapy (recent examples include [31 - 35]); often in these applications the model is combined with a tumor growth model, e.g. [31, 36, 37]. The LQ equations, and a wide range of ODE generalizations to approximate such additional aspects as cell cycle kinetics or cell population heterogeneity, are also now heavily used for analyzing cell killing in experimental radiobiology (recent surveys include [38, 39]). The generalizations, e.g. [40], are typically somewhat less practical in that they involve extra adjustable parameters.
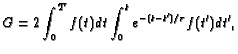 where
where