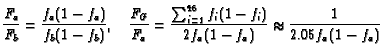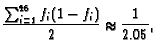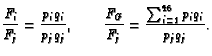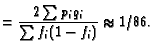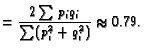Overview. The basic theoretical results on randomness in chromosome aberration production can be obtained by using contingency tables based on chromosome arm lengths (Savage and Papworth 1982). For the special, comparatively simple, case of two breaks, dominant at moderately low doses of sparsely ionizing radiations, the contingency table approach leads to various explicit equations, discussed below. In applications to high doses and/or complex aberrations, computer generalizations (Chen et al. 1995, 1996, 1997) of the explicit equations are needed, and the implications of randomness become model-dependent, e.g. are different for the breakage-and-reunion compared to the recombinational-repair model. The rest of this Appendix gives some details on the assumptions, consequences and empirical status of randomness, attempting inter alia to clear up some confusions which have arisen.
Randomness assumptions. It is sometimes useful to distinguish between randomness of DSB induction and randomness of misrejoining, related to the distinction between DSB allocation to regions in the genome and chromosome participation in aberrations (Savage 1991). Randomness of DSB induction would mean that just after an acute dose the average number of DSBs in any portion of the genome is proportional to the size (i.e. the genomic content, measured in Mbp). Averages over large DNA stretches, corresponding to the limit of resolution of the aberration assay, are meant; for example, whether randomness of DSB induction holds at the nucleosome level, for stretches as short as 200 bp, is not directly relevant in the usual aberration assays; even non-randomness in a substantial portion of a chromosome arm compared to other portions of that same arm, as suggested by some aberration experiments (Johnson et al. 1999, Xiao and Natarjan 1999a) and mutation experiments (e.g. Zhu et al. 1996), would usually not influence the frequencies scored in a FISH assay provided the average over the whole arm is representative of the genome as a whole.
Randomness of misrejoining would mean that the probability of DSB free ends interacting is the same regardless of which free ends are involved. Deviations from randomness of misrejoining occur due to proximity effects which bias for misrejoining of DSBs initially formed close together. Proximity effects bias for rings relative to simple interchanges (Savage and Papworth 1973), for insertions relative to S&S types 2F and 2G, and against complex interchanges relative to simple interchanges (Sachs et al. 1997a). By using interaction sites (Kellerer 1985, Savage 1996) these deviations from randomness can be systematically (though only approximately) incorporated into the calculations (Chen et al. 1996, 1997). Complete randomness of reactive DSB induction and misrejoining will now temporarily be assumed; later we outline changes in the predictions caused by taking proximity effects for misrejoining into account. Completeness will also be assumed.
Theoretical results for the case of one reactive DSB.
The simplest situation is when
aberration assays are used to estimate the number of breakpoints
(i.e. DSBs which misrejoin rather than restituting) in a particular
region of the genome. Randomness would mean that the average
number of breakpoints is directly proportional to the size
of the
region observed (review in Johnson et al. 1999). Specifically,
suppose in one experiment a portion of the genome is painted with a
FISH colour, the size relative to the size of the whole genome
being ![]() . Suppose in an otherwise identical experiment a
different fraction,
. Suppose in an otherwise identical experiment a
different fraction, ![]() , of the genome is painted. Then
randomness predicts the following simple proportionalities
for the observed breakpoint frequencies
, of the genome is painted. Then
randomness predicts the following simple proportionalities
for the observed breakpoint frequencies ![]() ,
, ![]() , and for the
frequency
, and for the
frequency ![]() of breakpoints in the whole genome:
of breakpoints in the whole genome:
Theoretical results for the case of two reactive DSBs. The most frequent applications of randomness in the literature are to exchanges involving two and only two DSBs in situations where more complicated interactions are negligible. The basics are the following.
Symmetric and Asymmetric Aberrations. In this special case of exactly two DSBs, randomness of misrejoining implies equality for the frequencies of `symmetric' and `asymmetric' aberrations (Savage and Papworth 1982), specifically
| (3) |
For translocations observed in one-colour painting, equations corresponding to Eq. 1 are (Lucas et al. 1992)
Eq. 4 also applies to dicentrics in the case where one
homologue pair is painted, provided the convention is made that a
dicentric involving both homologues is counted as contributing once
to ![]() but not contributing to
but not contributing to ![]() at all.
at all.
The reasoning behind Eq. 4 is the following (Savage and
Papworth 1982, Lucas et al. 1992). With a fraction ![]() of the genome painted, a fraction
of the genome painted, a fraction ![]() is counterstained. For
two DSBs, an observed interchange (i.e. an interchange involving a junction
between FISH paint and counterstain) requires that one of the two
DSBs is on the painted portion (probability
is counterstained. For
two DSBs, an observed interchange (i.e. an interchange involving a junction
between FISH paint and counterstain) requires that one of the two
DSBs is on the painted portion (probability ![]() ) and the other
DSB is on the counterstained portion (probability
) and the other
DSB is on the counterstained portion (probability ![]() ). This
argument gives the equation for
). This
argument gives the equation for ![]() . To get the formula
for
. To get the formula
for ![]() , similar reasoning is applied to the chromosomes one at a
time (Lucas et al. 1992), taking into account that each
interchange involves two different chromosomes.
, similar reasoning is applied to the chromosomes one at a
time (Lucas et al. 1992), taking into account that each
interchange involves two different chromosomes.
An Example. For example, each homologue of human
chromosome 2 is about 255 Mbp in size, while the total for all 46
chromosomes is about 6370 Mbp. Thus if one homologue
were painted,
![]() . In the more
realistic case of both homologues painted,
. In the more
realistic case of both homologues painted,
![]() . If
the homologue were painted the frequency of observed translocations
involving this homologue would, according to Eq. 4, be
approximately 7.9% of the frequency for translocations in the
entire genome, i.e.
. If
the homologue were painted the frequency of observed translocations
involving this homologue would, according to Eq. 4, be
approximately 7.9% of the frequency for translocations in the
entire genome, i.e.
| (7) |
Intrachanges. Formulae similar to Eq.
4, obtained by similar reasoning, hold for intrachanges
(rings and inversions) (Sachs et al. 1993, Wu et al. 1997b, Lucas
et al. 1999). We next state the results for rings but
by using Eq. 2 analogous equations are obtained for
inversions, or for the sum of rings and inversions. Let ![]() and
and
![]() denote the lengths of the short and long arms of the
denote the lengths of the short and long arms of the
![]() chromosome. Thus
chromosome. Thus
![]() and for humans
and for humans
![]() .
Then for centric rings, with
.
Then for centric rings, with ![]() the frequency for the
the frequency for the ![]() chromosome and
chromosome and ![]() that for the whole genome, one has
that for the whole genome, one has
| (9) |
In the case under consideration, where there are just two DSBs, the arguments sketched above can be extended to a randomness prediction of the frequency for centric rings compared to dicentrics (or, equivalently by Eq. 2, pericentric inversions compared to translocations); for example for the whole genome randomness predicts the frequency ratio (Savage and Papworth 1973; Hlatky et al. 1992)
It is well known, however, that there are actually far more centric rings relative to dicentrics than Eq. 10 predicts (Savage and Papworth 1973, Hlatky et al. 1992), due to proximity effects.
Proximity effects and interaction sites. Proximity
effects can typically be incorporated into a model by using
`sites' or by using `interaction distances' (Kellerer 1985).
Site models are presumably less realistic than
interaction distance models but typically involve fewer adjustable
parameters. Conceptually, sites are subregions of the cell nucleus
within which interactions can take place (survey in Savage 1996).
For example, CAS incorporates proximity effects quantitatively
by dividing the nucleus into some number ![]() of sites, with
interactions possible only between reactive DSBs
in the same site (see the Methods section above).
For whole-genome scoring, Eq. 10 is replaced in a
site model, approximately, by
of sites, with
interactions possible only between reactive DSBs
in the same site (see the Methods section above).
For whole-genome scoring, Eq. 10 is replaced in a
site model, approximately, by
Theoretical results for more than two DSBs: computer extensions of the equations. At doses of several Gy or higher, exchanges involving three or more breaks occur with significant frequencies. Then the above equations require some corrections, and in addition results for visibly complex aberrations are needed. Even for three breaks explicit equations similar to equation 4, or contingency tables, though putatively applicable, become enmeshed in a very complicated welter of contingencies. To make matters worse, the number of reactive breaks in an interaction site is not fixed at two or three or any given number. For low LET it is presumably governed by a Poisson distribution whose average is proportional to the genomic content of the site, and for high LET more complicated statistical distributions are needed (Kellerer 1985, Chen et al. 1997). In addition, the implications of randomness are in general model-dependent. Assuming doses so low that the frequency of complex aberrations is negligible, the results above (e.g. Eqs. 2 - 4, 8, 10, and 11) are valid no matter whether we assume the random-breakage-and-reunion model, the random recombinational repair model, or a random Revell-type exchange model. However, at higher doses, the predictions of the different models diverge.
Because of all these complications, explicit equations are problematical at higher doses. The only explicit equation currently known is that under the breakage-and-reunion model one has the following generalization of part of Eq. 2 (Lucas et al. 1996):
To indicate the magnitude of the corrections to explicit equations involved at higher doses, Figure 5 shows the fraction of apparently simple interchanges predicted for each chromosome by Lucas' chromosome participation formula 4 and by the random breakage-and-reunion model, at low, intermediate and high doses. It is seen that for low doses, the formula is accurate, but at 9 Gy the actual model gives significant deviations from the formula approximating the model. Even at 3 Gy there are perceptible deviations; these deviations are in the same direction as, but much smaller than, the experimental deviations seen by Barquinero et al., i.e. the actual model result calculated by CAS is slightly but not very significantly closer to experiment than the Lucas approximation.
Experimental results on randomness. There is strong evidence that various predictions of randomness modulated by proximity hold, at least within 50% or better (e.g. Savage 1976, Chadwick and Leenhouts 1981, Savage and Papworth 1982, Savage 1991, Kovacs et al. 1994, Finnon et al. 1995, Muhlmann-Diaz and Bedford 1995, Lucas et al. 1996, Chen et al. 1996, 1997, Sachs et al. 1997a, Boei et al. 1998a, Johnson et al. 1998, 1999). The predictions which have been found to hold approximately include Eqs. 1, 2, 4, 8, 11, their corrections by computer simulations at moderately large doses, Eq. 12, and various additional results, also obtained by computer simulations, for visibly complex aberrations (e.g. Table 3 and Fig. 3B).
However, there is also considerable evidence that some deviations from the randomness predictions may occur. As regards Eq. 12, which is the most clear-cut case because equality is expected even at doses so high that a significant fraction of aberrations are complex (i.e. even when distortion effects are significant), there have been repeated reports that an excess of apparently simple translocations is observed, with ratios of apparently simple translocations to apparently simple dicentrics ranging from about 1 to about 2. For example, the two recent, very large, data sets used in the main text above (Barquinero et al. 1998, Simpson and Savage 1996), both do show some excess (Tables 1 and 5), though by much less than the factor of about 2 found in some earlier, smaller data sets.
In addition the Lucas chromosome participation formula, Eq. 4, underpredicts, somewhat, observed apparently simple aberrations involving most small chromosomes, and correspondingly overpredicts apparently simple aberrations involving most large chromosomes, for some large data sets on human lymphocytes (Knehr et al. 1996, Barquinero et al. 1998, Cigarran et al. 1998). More precise CAS calculations give at 3 Gy results basically similar to the Lucas formula (Fig. 5), though marginally closer to the lymphocyte data (Table 5). The present analysis confirms a substantial tendency for Chr. 3 to underparticipate, as compared to expectations based on randomness, not only in apparently simple aberrations, as pointed out by Barquinero et al., but also in visibly complex ones (Fig. 3, Table 5). For the fibroblast data no clear trend for large chromosomes to underparticipate in aberrations is present (Table 1). Many other possible deviations from randomness have been discussed (e.g. Xiao and Natarjan 1999b), but at present (apart perhaps from hot spots near centromeres, telomeres, and intrachromosomal telomeric sequences) do not appear to constitute systematic patterns applicable to different data sets.
Intra-arm intrachanges. Comparatively little is known experimentally about intra-arm intrachanges (i.e. paracentric inversions and acentric rings). Much as in Eq. 10 above, randomness also predicts various intrachange frequency ratios, e.g.
References
BALLARINI F; MERZAGORA M; MONFORTI F; DURANTE M; GIALANELLA G; GROSSI GF; PUGLIESE M; OTTOLENGHI A., 1999. Chromosome aberrations induced by light ions: Monte Carlo simulations based on a mechanistic model. International Journal of Radiation Biology, 75:35-46.
BARQUINERO JF; KNEHR S; BRASELMANN H; FIGEL M; BAUCHINGER M., 1998. DNA-proportional distribution of radiation-induced chromosome aberrations analysed by fluorescence in situ hybridization painting of all chromosomes of a human female karyotype. International Journal of Radiation Biology, 74:315-23.
BAUCHINGER, M. and SCHMID, E., 1998. LET dependence of yield ratios of radiation-induced intra- and interchromosomal aberrations in human lymphocytes. International Journal of Radiation Biology, 74, 17-25.
BOEI JJ; VERMEULEN S; NATARAJAN AT, 1998a. Dose-response curves for X-ray induced interchanges and interarm intrachanges in human lymphocytes using arm-specific probes for chromosome 1. Mutat Res, 404:45-53.
BOEI, J.J. and NATARAJAN, A.T., 1998. Combined use of chromosome painting and telomere detection to analyse radiation-induced chromosomal aberrations in mouse splenocytes. International Journal of Radiation Biology, 73, 125-33.
BOEI, J.J., VERMEULEN S., FOMINA, J. and NATARAJAN, A.T., 1998b. Detection of incomplete exchanges and interstitial fragments in X-irradiated human lymphocytes using a telomeric PNA probe. International Journal of Radiation Biology, 73, 599-603.
BRENNER DJ, LUBIN JH and RON E., 1998. Moving from under the lamppost: can epidemiologists and radiobiologists work together? Nuclear Energy, 37, 25-31.
CHADWICK, K.H. and LEENHOUTS, H.P. 1981. The molecular theory of radiation biology. New York, Springer-Verlag, pp. 62-63 and 126-132.
CHADWICK, K.H. and LEENHOUTS, H.P. 1998. Radiation induced chromosome aberrations: some biophysical considerations. Mutat Res, 404, 113-7.
CHATTERJEE, A. and HOLLEY, W.R., 1991, Energy deposition mechanisms and biochemical aspects of DNA strand breaks by ionizing radiation, Int. J. Quant. Chem. 391, 709-727.
CHEN, A.M., LUCAS, J.N., HILL, F.S., BRENNER, D.J. and SACHS, R.K. 1995. Chromosomal aberrations produced by ionizing radiation: Monte Carlo simulations and chromosome painting data. Computer Applications in the Biosciences (CABIOS), 11, 389-397.
CHEN A.M., LUCAS J.N., BRENNER, D.J., HILL, F.S. and SACHS, R.K., 1996. Proximity effects for chromosome aberrations measured by FISH. International Journal of Radiation Biology, 69 411-420.
CHEN, A.M., LUCAS, J.N., SIMPSON, P.J., GRIFFIN, C.S., SAVAGE,
J.R.K., BRENNER, D.J., HLATKY, L.R. and SACHS, R.K., 1997.
Computer simulation of FISH data on chromosome aberrations
produced by X-rays or ![]() particles. Radiation
Research, 148 S93-S101.
particles. Radiation
Research, 148 S93-S101.
CHEN, A.M., HAHNFELDT, P. and SACHS, R.K., 1998. Chromosome
aberration simulator (CAS). User's manual, source code and
executables for UNIX or Windows95/98 available on request or via
anonymous ftp at math.berkeley.edu (directory
pub/Preprints/Rainer_Sachs/CAS) or from URL
/![]() chen/.
chen/.
CIGARRAN S; BARRIOS L; BARQUINERO JF; CABALLIN MR; RIBAS M; EGOZCUE J., 1998. Relationship between the DNA content of human chromosomes and their involvement in radiation-induced structural aberrations, analysed by painting. International Journal of Radiation Biology, 74:449-55.
CORNFORTH M. N. and BEDFORD J. 1993, Ionizing radiation damage and its early development in chromosomes. Advances in radiation biology, vol. 17. DNA and chromatin damage caused by radiation. Edited by: J. T. Lett and W. K. Sinclair (San Diego, Academic press), 423-496.
CORNFORTH M.N. 1998, Radiation-induced damage and the formation of chromosomal aberrations. In: DNA damage and repair, J.A. Nickoloff and M.F. Hoekstra eds. (Totowa, N.J., Humana Press), 559-585.
CREMER T., KURZ A., ZIRBEL R., DIETZEL S., RINKE B., SCHROCK E., SPEICHER M. R., MATHIEU U., JAUCH A., EMMERICH P., SCHERTAN H., RIED T., CREMER C. and LICHTER P. 1993, Role of chromosome territories in the functional compartmentalization of the cell nucleus. Cold Spring Harbor Symposia on Quantitative Biology, 58, 777-792.
EDWARDS, A.A., MOISEENKO, V.V. and NIKJOO, H., 1996. On the mechanism of the formation of chromosomal aberrations by ionizing radiation. Radiation and Environmental Biophysics, 35, 25-30.
EDWARDS AA., 1997. The use of chromosomal aberrations in human lymphocytes for biological dosimetry. Radiation Research, 148:S39-44.
EDWARDS, A.A., and SAVAGE J.R.K. 1999. Is there a simple answer to the origin of complex exchanges? Int. J. Radiat. Biol., 00, 000-000.
FINNON P., LLOYD D.C., EDWARDS A.A., 1995. Fluorescence in situ hybridization detection of chromosomal aberrations in human lymphocytes: applicability to biological dosimetry. International Journal of Radiation Biology, 68:429-35.
GOODHEAD D. T., THACKER J. and COX R. 1993, Weiss Lecture. Effects of radiations of different qualities on cells: molecular mechanisms of damage and repair. International Journal of Radiation Biology, 63, 543-56.
GOODHEAD, D.T., 1987, Relationship of microdosimetric techniques to applications in biological systems. In: The Dosimetry of ionizing radiation, Vol. II. Edited by: K. Kase, B. Bjarngard and F. Attix (Academic Press, Orlando), pp. 1-89.
GRIFFIN, C.S., MARSDEN, S.J., STEVENS, D.L., SIMPSON, P. and SAVAGE, J.R. 1995. Frequencies of complex chromosome exchange aberrations induced by 238Pu alpha-particles and detected by fluorescence in situ hybridization using single chromosome-specific probes. Int. J. Radiat. Biol., 67, 431-439.
GRIFFIN, C.S., HILL, M.A., PAPWORTH, D.G., TOWNSEND, K.M., SAVAGE, J.R. and GOODHEAD, D.T. 1998. Effectiveness of 0.28 keV carbon K ultrasoft X-rays at producing simple and complex chromosome exchanges in human fibroblasts in vitro detected using FISH. International Journal of Radiation Biology, 73, 591-8.
HLATKY, L.R., SACHS, R.K., and HAHNFELDT, P., 1992. The ratio of centric rings to dicentrics in irradiated human lymphocytes. Radiation Research, 129, 304-308.
JEGGO, P.A., 1998. Identification of genes involved in repair of DNA double-strand breaks in mammalian cells. Radiation Research, 150, S80-91.
JOHNSON, K.L., TUCKER J.D., and NATH J. 1998. Frequency, distribution and clonality of chromosome damage in human lymphocytes by multi-color FISH. Mutagenesis, 13, 217-27.
JOHNSON, K.L., BRENNER D.J., NATH, J., TUCKER, J.D., and GEARD,
C.R. 1999. Radiation-induced misrejoined breakpoints in human
chromosomes: random or non-random.
textitRadiation Research,
000, 000-000.
KELLERER, A.M., 1985, Fundamentals of Microdosimetry. In: The Dosimetry of Ionizing Radiation, Vol. I. Edited by: K. Kase, B. Bjarngard and F. Attix (Academic Press, Orlando), pp. 77-162.
KNEHR, S., ZITZELSBERGER, H., BRASELMANN, H., NAHRSTEDT, U. and BAUCHINGER, M., 1996. Chromosome analysis by fluorescence in situ hybridization: further indications for a non-DNA-proportional involvement of single chromosomes in radiation-induced structural aberrations. International Journal of Radiation Biology 70, 385-392.
KNEHR, S., ZITZELSBERGER, H., NAHRSTEDT, U. and BAUCHINGER, M., 1998. FISH-based analysis of radiation-induced chromosomal aberrations using different nomenclature systems. International Journal of Radiation Biology, 73, 135-41.
KODAMA, Y., NAKANO, M., OHTAKI, K., AWA, A.A., NAKAMURA, N. and DELONGCHAMP, R. 1997. Estimation of minimal size of translocated chromosome segments detectable by fluorescence in situ hybridization Int. J. Radiat. Biol. 71, 35-39.
KOVACS M. S., EVANS J. W., JOHNSTONE I. M. and BROWN J. M. 1994, Radiation-induced damage, repair and exchange formation in different chromosomes of human fibroblasts determined by fluorescence in situ hybridization. Radiation Research, 137, 34-43.
LINDHOLM, C., LUOMAHAARA, S., KOIVISTOINEN, A., ILUS, T., EDWARDS, A.A., SALOMAA, S. 1998. Comparison of dose-response curves for chromosomal aberrations established by chromosome painting and conventional analysis. International Journal of Radiation Biology, 74, 27-34.
LUCAS J. N., AWA A., STRAUME T., POGGENSEE M., KODAMA Y., NAKANO M., OHTAKI K., WEIER H. U., PINKEL D., GRAY J. and LITTLEFIELD G., 1992. Rapid translocation frequency analysis in humans decades after exposure to ionizing radiation. International Journal of Radiation Biology, 62, 53-63.
LUCAS J.N. and SACHS R.K., 1993. Using 3-colour chromosome painting to decide between chromosome aberration models. Proceedings of the National Academy of Sciences, USA, 90, 1484-1487.
LUCAS J.N., CHEN A.M. and SACHS R. K., 1996. Theoretical predictions on equality of radiation-produced dicentrics and translocations detected by chromosome painting. International Journal of Radiation Biology, 69, 145-153.
LUCAS, JN; DENG, W; ORAM, SW; HILL, FS; DURANTE, M., GEORGE, K., WU, H, AND YANG, T., 1999. Theoretical and experimental tests of a chromosomal fingerprint for densely ionizing radiation based on F ratios calculated from stable and unstable chromosome aberrations. Radiation Research, 151:85-91.
NORMAN A; SASAKI MS., 1966. Chromosome-exchange aberrations in human lymphocytes. International Journal of Radiation Biology and Related Studiesin Physics, Chemistry and Medicine, 11:321-8.
PHILLIPS JW; MORGAN WF. Illegitimate recombination induced by DNA double-strand breaks in a mammalian chromosome, 1994. Molecular and Cellular Biology, 14:5794-803.
RADIVOYEVITCH, T., 1997. Mathematical models of misrejoining DNA double-strand breaks and DNA fragment size distributions, PhD Thesis, Department of Biometry and Epidemiology, Medical University of South Carolina, Charleston, SC 29425.
RADIVOYEVITCH, T., HOEL, D.G., CHEN, A. and SACHS, R.K., 1998. Misrejoining of double-strand breaks after X irradiation: relating moderate to very high doses by a Markov model. Radiation Research, 149, 59-67.
RESNICK, M.A., BENNET, C., KOUPRINA, N, LARIONOV, V., LEWIS. K., TRAN, H., 1995. Recombinational repair of double strand breaks: twenty years later. In: Radiation Research 1895-1995, U. Hagen, D. Harder, H. Jung, and C. Streffer eds., H. Stuertz, Wuerzberg, pp. 362-367.
REVELL S.H., 1974. The breakage-and-reunion theory and the exchange theory for chromosomal aberrations induced by ionizing radiation: A short history. Advances in Radiation Biology 4, 367-416.
ROOS H; SCHMID E, 1998. Analysis of chromosome aberrations in human peripheral lymphocytes induced by 5.4 keV x-rays. Radiation and Environmental Biophysics, 36:251-4.
SACHS R. K., AWA A., KODAMA Y., NAKANO M., OHTAKI K. and LUCAS J. N. 1993, Ratios of radiation-produced chromosome aberrations as indicators of large-scale DNA geometry during interphase. Radiation Research, 133, 345-350.
SACHS, R.K., CHEN, A.M., and BRENNER, D.J., 1997a. Review: proximity effects in the production of chromosome aberrations by ionizing radiation. Int. J. Radiat. Biol., 71, 1-19.
SACHS, R.K., BRENNER, D.J., CHEN, A.M., HAHNFELDT, P. and HLATKY, L.R., 1997b. Intra-Arm and Inter-Arm Chromosome Intrachanges: Tools for Probing Chromatin Geometry and Dynamics, Radiation Research, 148, , 330-339.
SACHS, R.K., HAHNFELDT, P. and BRENNER, D.J., 1997c. Review: The link between low-LET dose-response relations and the underlying kinetics of damage production/repair/misrepair. Int. J. Radiat. Biol. 72, 351-374.
SACHS, R.K., BRENNER, D.J., HAHNFELDT, P.J. and HLATKY, L.R., 1998. A Formalism for Analyzing Large-Scale Clustering of Radiation-Induced Breaks Along Chromosomes. International Journal of Radiation Biology, 74, 185-206.
SACHS, R.K., A.M. CHEN, P.J. SIMPSON, L.R. HLATKY, P. HAHNFELDT, and J.R.K. SAVAGE, 1999. Clustering of radiation-produced breaks along chromosomes: modeling the effects on chromosome aberrations International Journal of Radiation Biology, 00, 000-000.
SAVAGE JR; PAPWORTH DG, 1969. Distortion hypothesis: an alternative to a limited number of sites for radiation-induced chromosome exchange. Journal of Theoretical Biology, 22:493-514.
SAVAGE J. R. K. and PAPWORTH D. G. 1973, The relationship of radiation-induced yield to chromosome arm number. Mutation Research, 19, 139-143.
SAVAGE, J.R., 1976. Classification and relationships of induced chromosomal structural changes. Journal of Medical Genetics, 1976, 103-22.
SAVAGE J. R. K. and PAPWORTH D. G. 1982, Frequency and distribution studies of asymmetrical versus symmetrical chromosome aberrations. Mutation Research, 95, 7-18.
SAVAGE J.R., 1991. Testing the participation of chromosomes in structural aberrations. In: Eukaryotic chromosomes : structural and functional aspects editors, R.C. Sobti, G. Obe. Berlin; Springer-Verlag.
SAVAGE J. R. K. and SIMPSON P. 1994, FISH "painting" patterns resulting from complex exchanges. Mutation Research, 312, 51-60.
SAVAGE, J.R. and TUCKER, J.D. 1996. Nomenclature systems for FISH-painted chromosome aberrations. Mutation Research, 366, 153-161.
SAVAGE, J.R. 1996. Insight into sites. Mutation Research, 366, 81-95.
SAVAGE, J.R. 1998. A brief survey of aberration origin theories. Mutat Res,404, 139-47.
SAVAGE J. R. K. and PAPWORTH D. G. 1998. An investigation of LET "fingerprints" in Tradescantia. Mutation Research, 422, 313-322.
SIMPSON P.J. and SAVAGE J.R.K., 1996. Dose-response curves for simple and complex chromosome aberrations induced by x-rays and detected using fluorescence in situ hybridization. International Journal of Radiation Biology 69, 429-436.
SIMPSON P.J., PAPWORTH D.G. and SAVAGE J.R.K., 1999. X-ray induced simple, pseudosimple, and complex exchanges involving two distinctly painted chromosomes. International Journal of Radiation Biology 000, 000-000.
TUCKER JD; EASTMOND DA; LITTLEFIELD LG., 1997. Cytogenetic end-points as biological dosimeters and predictors of risk in epidemiological studies. Iarc Scientific Publications, 142, 185-200.
WU H; DURANTE M; GEORGE K; YANG TC, 1997a. Induction of chromosome aberrations in human cells by charged particles. Radiation Research, 148:S102-7.
WU, H., DURANTE, M., SACHS, R.K. and YANG, T.C. 1997b. A random walk model for interphase chromosomes and the relationship between centric rings, acentric rings and excess acentric fragments induced by low-LET radiation. Int. J. Radiat. Biol., 71, 487-496.
WU, H., GEORGE, K., and YANG, T.C. 1998. Estimate of true incomplete exchanges using fluorescence in situ hybridization with telomere probes. Int. J. Radiat. Biol., 73, 521-7.
XIAO, Y., and NATARAJAN, A.T., 1999a. The heterogeneity of Chinese hamster X chromosome in X-ray induced chromosomal aberrations. Int. J. Radiat. Biol., 00, 000-000.
XIAO, Y., and NATARAJAN, A.T., 1999b. Non-random involvement of Chinese hamster chromosomes 3, 4, 8, and 9 in X-ray induced chromosomal aberrations. Int. J. Radiat. Biol., 00, 000-000.
ZHU LX; WALDREN CA; VANNIAS D; HEI TK. Cellular and molecular analysis of mutagenesis induced by charged particles of defined linear energy transfer. Radiation Research, 145:251-9, 1996.
Figure 1: Two models for a visibly complex
interchange. Panel A shows three chromosomes; one chromosome is
painted with a FISH colour (shown black); the other two are
counterstained (shown white). Centromeres, here shown as circles, are
assumed recognizable (e.g. with a pancentromeric stain) throughout
the analysis.
Three DSBs are shown; for example the first DSB has free ends ![]() and
and ![]() . All DSBs shown are assumed to be reactive, i.e. capable
of misrejoining. According to the
breakage-and-reunion model, one possible exchange is for the DSB
free ends to misrejoin in the reaction [
. All DSBs shown are assumed to be reactive, i.e. capable
of misrejoining. According to the
breakage-and-reunion model, one possible exchange is for the DSB
free ends to misrejoin in the reaction [
![]() ];
this is a cycle of order 3 (Sachs et al. 1999).
This reaction produces the final pattern
shown in panel B. In a FISH experiment, only the two components
showing a FISH signal are scored. Even so, the interchange
recognizably involves at least 3 DSBs, i.e. the pattern
is visibly complex,
because the number of centromeres on the two scored components is
wrong for a complete reciprocal exchange involving just two DSBs. The
particular pattern in panel B, with two bicoloured final components
having three centromeres, is designated 2G in the S&S
aberration classification system (Savage and Simpson 1994).
];
this is a cycle of order 3 (Sachs et al. 1999).
This reaction produces the final pattern
shown in panel B. In a FISH experiment, only the two components
showing a FISH signal are scored. Even so, the interchange
recognizably involves at least 3 DSBs, i.e. the pattern
is visibly complex,
because the number of centromeres on the two scored components is
wrong for a complete reciprocal exchange involving just two DSBs. The
particular pattern in panel B, with two bicoloured final components
having three centromeres, is designated 2G in the S&S
aberration classification system (Savage and Simpson 1994).
The same 2G observed pattern can be made according to the
recombinational-repair mechanism, but more breaks are involved.
The next two panels give an example. Panel C shows two
radiation-induced DSBs, ![]() and
and ![]() . Suppose the first DSB,
. Suppose the first DSB,
![]() , `attacks' a third, counterstained, chromosome at the
otherwise undamaged location
, `attacks' a third, counterstained, chromosome at the
otherwise undamaged location ![]() and a complete reciprocal
exchange [
and a complete reciprocal
exchange [
![]() ], occurs; suppose in addition
], occurs; suppose in addition ![]() attacks on the same arm, further from the centromere, and the
complete reciprocal exchange [
attacks on the same arm, further from the centromere, and the
complete reciprocal exchange [
![]() ] occurs. The
outcome of these exchanges
is shown in panel D. The sizes of some of the final
components are different than in panel B, but the observed
aberration pattern is again 2G.
] occurs. The
outcome of these exchanges
is shown in panel D. The sizes of some of the final
components are different than in panel B, but the observed
aberration pattern is again 2G.
Figure 2: An incomplete pattern. The conventions are the same as in Figure 1. Panel A shows final aberration pattern 2F in the S&S classification. Because neither of the counterstained segments contains a centromere, the pattern is a visibly complex interchange - at least three DSBs must have been involved, and this fact is evident even if only final components which show a FISH signal are examined.
Panel B shows a pattern sometimes observed, designated incomplete type I (Simpson and Savage 1996) or pattern 2BI (Barquinero et al. 1998). The optimal reclassification of the pattern as a complete pattern is somewhat problematic. One plausible explanation is that the acentric painted fragment is actually attached to an acentric counterstained fragment, just as in panel A, but the counterstained fragment is too small to be seen. Then the S&S pattern would be 2F, as in panel A. However the observed pattern could possibly also indicate an incomplete version of S&S pattern 2B (Simpson and Savage 1996).
Figure 3: Scatter plots for predicted and observed chromosome participation. Observed values of the per-cell frequency of certain aberrations for particular chromosomes painted, compared to the values calculated with CAS using the breakage-and-reunion model and the parameters of Table 2. Panel A. Values for apparently simple interchanges in the SS4 data. The SS4 values are obtained from Table 1, dividing the `2A+2B' row by the `Cells' row. For apparently simple interchanges the CAS values are approximately equal to those calculated with the Lucas formula (Appendix). The line is the best-fitting straight line. From the values shown, a correlation coefficient of 0.92 (p=0.01) was calculated. This correlation coefficient, and other correlation coefficients for chromosome participation, are shown in Table 3. Panels B-D respectively show plots similar to that in panel A, but for TVC in Table 1, 2A+2B in Table 5, and TVC in Table 5 respectively.
Figure 4. Dose-response relations. Fibroblast data
(Simpson and Savage 1996) for apparently simple interchanges
(ASI=2A+2B, squares) and total visibly complex patterns
(TVC, diamonds) are shown; aberration frequencies are summed
for the 6 homologue pairs painted one pair at a time.
In panel A the parameters of Table 2 are used for the random
breakage-and-reunion model (BR, solid lines) and random
recombinational-repair model (RR, dotted lines). The latter is
quite innacurate for the visibly complex aberrations. If only
apparently simple interchanges are considered, the constraints on
the model parameters are relaxed (panel B);
then the optimal BR curve, obtained by
replacing ![]() =2.3 with
=2.3 with ![]() =2.15, is
quite close to the data.
=2.15, is
quite close to the data.
For apparently simple aberrations the BR model curves are quadratic only at low doses, and the RR curves are linear only at low doses. The downward curvatures seen at higher doses are mainly due to distortion, i.e. to the indirect effects of complex aberration formation (Savage and Papworth 1969); in the BR case saturation also plays a role (Chen et al. 1997). For aberrations which are simple rather than just apparently simple, distortion effects are even stronger (curves not shown).
Figure 5: Theoretical corrections to the Lucas chromosome participation formula at higher doses. Squares show the results of Eq. 4 for the relative frequency of observed apparently simple translocations that involve a particular homologue pair in a human female genome. 23 experiments with equal cell numbers and with one homologue pair painted in each experiment are assumed. The total is 200% because two differently numbered chromosomes are involved in each observed translocation. Corrections are needed for Eq. 4 at doses higher than a few Gy. CAS calculations, assuming the random breakage-and-reunion model with the B(ii) parameters of Table 2, are given for 1.5 Gy (light solid line), for 3 Gy (heavy solid line), and for 9 Gy (dotted line). The lines connect discrete points to guide the eye. The differences between the actual model predictions and the approximation given by Eq. 4 are due to distortion and saturation; they increase as the dose increases, and are more pronounced for truly simple translocations (curves not shown) than for the case shown here of apparently simple translocations.