


Next: Discussion
Up: ms
Previous: Methods
Preview. For the one-colour data sets, we first
apply the breakage-and-reunion model to the 4 Gy fibroblast
data (Simpson and Savage 1996), then the
recombinational-repair model. Thereafter somewhat smaller
fibroblast data sets, for 2 Gy and 6 Gy, and the very large
lymphocyte data set of Barquinero et al. (1998)
will be analyzed similarly.
Finally, other data, for multi-colour painting, is briefly discussed.
Fibroblast data.
Table 1 shows fibroblast data (Simpson and Savage 1996) at 4 Gy
for 6
different chromosome homologue pairs painted (effectively
one pair at a time), and for a number of different aberration types.
In the
table, the data is compared with the breakage-and-reunion model
predictions, calculated as described in the Methods section,
using the parameters 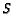 and
and 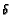 shown in Table 2.
It was found that only the apparently simple
centric rings depend sensitively on site number in the range
8-12 sites; predicted frequencies for the other aberration
categories listed remain roughly fixed if is
varied holding
shown in Table 2.
It was found that only the apparently simple
centric rings depend sensitively on site number in the range
8-12 sites; predicted frequencies for the other aberration
categories listed remain roughly fixed if is
varied holding
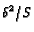 constant (i.e.
proportional to
constant (i.e.
proportional to 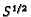 ).
).
[Insert Tables 1 and 2 and captions about here]
The participation of different chromosomes in aberrations was
further analyzed by determining correlation coefficients.
For example, in
Figure 3A, CAS model predictions for apparently simple interchanges
are shown in a scatter plot with the experimental results.
Figure 3B gives corresponding results for total visibly complex
aberrations. The correlation coefficients for these and other
aberration types are shown in Table 3. For the most part
strong correlations are seen.
[Insert Fig. 3 + table 3 + captions about here]
A similar analysis was carried out for the recombinational-repair
model. The best parameters for this model, considering all three
doses, were found to be 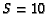 sites and =0.45/Gy (Table
2). Summary results, for totals over all chromosomes, are shown in
Table 4. It is seen that the model drastically underpredicts
aberration patterns 2F and 2G (Table 4). The model also
drastically underpredicts insertions, which are included among the
TVC category in Table 4. However, the chromosome participation
correlations remain high (Table 3).
sites and =0.45/Gy (Table
2). Summary results, for totals over all chromosomes, are shown in
Table 4. It is seen that the model drastically underpredicts
aberration patterns 2F and 2G (Table 4). The model also
drastically underpredicts insertions, which are included among the
TVC category in Table 4. However, the chromosome participation
correlations remain high (Table 3).
[Insert table 4 + caption about here]
Similar, but less detailed, analyses were carried out at 2 Gy and at
6 Gy, using the same parameters. The main results are shown in
Tables 3 and 4. It is again seen that the random
breakage-and-reunion model gives reasonably accurate results, that
the correlations are high, and that the recombinational-repair model
drastically underpredicts 2F and 2G patterns. One additional
detail of interest is that at 6 Gy, for very complex aberrations
involving 6 or more breaks, the observed:predicted numbers for the
breakage-and-reunion model are 38:10, i.e. there is substantial
underprediction of very complex aberrations, as was previously
also found at 4 Gy (Chen et al. 1997).
Theoretical dose-response curves for simple and complex
aberrations, incorporating the effects of
distortion and saturation, are compared to data at three doses
in Figure 4.
The data of Barquinero et al.
This experiment has results for all 23 homologue
pairs, painted one pair at a time.
Table 5 gives the experimental data, with one-way exchange
types I-VI reallocated via method (ii), described in the Methods
section. The table also gives the predictions of the
breakage-and-reunion model with the parameters shown in Table 2.
Table 3 gives some correlations for chromosome participation.
The breakage-and-reunion model predictions are reasonably accurate.
Their weakest feature is a systematic tendency to overpredict
visibly complex aberrations other than 2F and 2G patterns
(contrasting with the underpredictions found for the fibroblast
data in this category).
Some corresponding results for the recombinational-repair model
are given in Tables 3 and 4. Once more it is seen that
the recombinational-repair model drastically underpredicts 2F and
2G frequencies. For method (i) of reclassifying
apparently incomplete aberration patterns, discussed in the
Methods section, the predictions for 2F and 2G are somewhat
improved, being about 1/5 of the observed values
rather than about 1/10, but for method (iii) the discrepancy
is somewhat worse (details not shown).
[Insert Table 5 and caption about here]
Multicolour painting. Assuming completeness and
negligible DSB clustering along chromosomes,
the recombinational-repair
pathway cannot lead to an odd number of junctions between any
two colours, since each of its complete reciprocal exchanges makes
either two or zero colour junctions; the
breakage-and-reunion model, however, allows odd numbers of
colour junctions. In view of the theorem that for one FISH color
all patterns allowed by the breakage-and-reunion model are also
allowed by the recombinational-repair model (Sachs et al. 1999)
distinguishing between the models by looking for odd numbers of
color junctions is relevant only
for painting with two or more FISH colours.
Three low-LET two-colour data sets give the
following respective numbers of patterns recorded as having
just one junction between the two FISH colours
(rather than having an even number):
Lucas and Sachs (1993), 18; Chen et al. (1995), 3;
Simpson et al. (1999), 14. A three-colour data set (Knehr et al.
1999) gives additional examples, including some for low LET
irradiation.
Next: Discussion
Up: ms
Previous: Methods
Rainer &
1999-05-22