
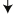 Review
Review
|
|
|
|
|
Review
|
Radiation-induced chromosome aberrations: insights gained from biophysical modeling
Lynn Hlatky, Dana Farber Cancer Institute, Harvard Medical School
Rainer K. Sachs, University of California Berkeley
Mariel Vazquez, University of California Berkeley
Michael N. Cornforth, University of Texas Medical Branch, Galveston
Summary
Enzymatic misrepair of ionizing-radiation induced DNA damage can produce large-scale rearrangements of the genome, such as translocations and dicentrics. These and other chromosome exchange aberrations can cause major phenotypic alterations, including cell death, mutation and neoplasia. Exchange formation requires that two (or more) genomic loci come together spatially. Consequently, the surprisingly rich aberration spectra uncovered by recently developed techniques, when combined with biophysically-based computer modeling, help characterize large-scale chromatin architecture in the interphase nucleus. Most results are consistent with a picture whereby chromosomes are mainly confined to territories, chromatin motion is limited, and interchromosomal interactions involve mainly territory surfaces. Aberration spectra and modeling also help characterize DNA repair/misrepair mechanisms. Quantitative results for mammalian cells are best described by a breakage-and-reunion model, suggesting that the dominant recombinational mechanism during the G0/G1 phase of the cell cycle is non-homologous end-joining of radiogenic DNA double strand breaks. In turn, better mechanistic and quantitative understanding of aberration formation gives new insights into health-related applications.
Key phrases: chromosome aberrations; ionizing radiation; chromosome territories; non-homologous end-joining; computer models.
|
Abbreviations: DSB, DNA double strand break; FISH, fluorescence in situ hybridization; mFISH, multiplex- (multicolor-, multifluor-) FISH; Gy, Gray (a unit of radiation dose; 1 Joule per kilogram). |
Introduction
Ionizing radiation produces rearrangements of the genome. When irradiation occurs during the G0/G1 phase of the cell cycle, large-scale rearrangements appear as exchange-type chromosome aberrations at the next mitosis. Such aberrations can alter cellular phenotypes, and are important in various areas of biology:
Here we review how data derived using recently developed cytogenetic approaches has been combined with biophysical modeling to elucidate the mechanisms and implications of aberration formation.
A colorful diversity of aberration types
Until rather recently, it was usually assumed that virtually all chromosome exchanges are simple, i.e. involve only two chromosome breaks (Fig. 1). However, chromosome "painting" techniques have now shown that complex aberrations, involving more than two breaks in a single configuration, are common.[Savage, 2000] Fig. 2 (modified from[Loucas, 2001] with permission) gives an example. Many whole-chromosome painting techniques are based on FISH.[Pinkel, 1986] More recent and sophisticated painting techniques, such as mFISH (Fig. 2) or spectral karyotyping, employ combinatorial hybridization schemes, allowing recognition of most exchanges between heterologous chromosomes.[Schrock, 2000; Cornforth, 2001; Greulich, 2000; Speicher, 1996; Lee, 2001] Still further extensions of this approach allow better recognition of exchanges between homologous chromosomes, better localization of exchange breakpoints within a chromosome, and better recognition of inversions (Fig. 1Bi).[Fauth, 2001; Saracoglu, 2001; Karhu, 2001; Wiegant, 2000] The intricate aberration spectra uncovered by chromosome painting give extra information about the mechanisms and geometric aspects of radiation damage.

Figure 1.
Simple chromosome aberrations. Constrictions represent centromeres; gaps indicate chromosome breaks that are caused either by prompt radiation damage or (in some cases according to some models) by subsequent enzymatic action. Simple aberrations, by definition, involve only two breaks. A. Two painted chromosomes, each of which contains a break. The result can be one of the following. i. A simple (reciprocal) translocation. While a cell harboring this type of aberration usually remains clonogenically viable, the resulting large-scale rearrangement of the genome can lead to phenotypic alterations, including mutation and oncogenic transformation. ii. A dicentric together with its associated acentric fragment. Dicentrics are usually clonogenically lethal because of segregation problems that arise during anaphase of mitosis. iii. A double restitution. Restitution is defined as rejoining in the pre-break configuration with, at worst, local alteration of the genome at the nucleotide level, such as a point mutation. B: A single chromosome with two breaks. Misrejoining can give simple intrachromosomal aberrations as follows. i. A (pericentric) inversion. ii. A centric ring with accompanying acentric fragment.As far as phenotypic changes are concerned, translocations and large inversions are expected to have similar consequences; likewise centric rings and dicentrics are comparable. Because they are often difficult to detect, inversions are among the least well-understood of the simple aberrations. Nevertheless, they are rather common, due to the proximity effects discussed in the text. The remaining types of simple aberrations, acentric rings and paracentric inversions (not shown), are also common but difficult to detect.
Figure 2.
An mFISH image for a chromosome spread containing both simple and complex exchanges. The cell, a human peripheral blood lymphocyte, was irradiated with a 4 Gy dose of gamma rays. In mFISH, chromosomes are assigned colors, corresponding to unique combinatorial hybridization signals that represent the 24 different types of chromosomes in the human genome. Exchange-type aberrations here include a complex rearrangement that simultaneously involves chromosomes 1, 2, 3, 9, 11 and 20 (white arrows). Two simple exchanges were also found in this cell, a dicentric involving chromosomes 1 and X (red arrows; one points to a chromosome near the limit of resolution) and a translocation involving chromosomes 12 and 21 (yellow arrows).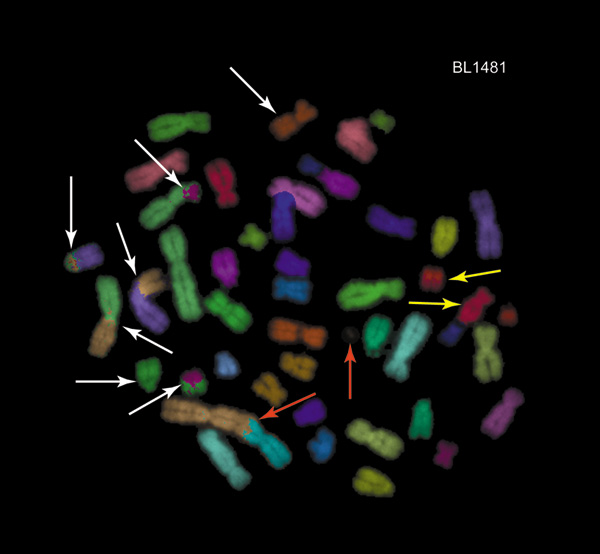
Aberration formation pathways
For over half a century chromosome aberrations have been extensively studied with regard to the repair/misrepair pathways involved in their formation.[Savage, 1998] The question as to what constitutes the principal underlying mechanism has remained controversial, sometimes contentiously so. Proposed models differ substantially in their predictions for the dose- and dose-rate dependency of aberration frequencies, and differ also with respect to the predicted spectra of aberrations
Debates have centered around three pathways, though these could be acting in parallel. Updated versions of the pathways are shown schematically in Fig. 3. The breakage-and-reunion pathway (Fig. 3A) corresponds, at the molecular level, to non-homologous end-joining.[Rothkamm, 2001; Hoeijmakers, 2001; Pfeiffer, 2000#188] An alternative scenario embraces a "1-hit" paradigm (Fig. 3B), in which a single radiation-induced DSB is sufficient to initiate an exchange with an otherwise undamaged portion of the genome, by interacting with a second, enzymatically-induced break.[Goodhead, 1993; Cucinotta, 2000] This scenario can correspond to homologous DNA repair,[Rothkamm, 2001; Hoeijmakers, 2001] a correspondence particularly evident if the interaction involves, as has sometimes been observed, limited sequence homology shared between two sites, such as that which can be provided by the abundance of repetitive DNA elements.[Thacker, 1999; Bode, 2000] The pathway shown in panel B has been termed recombinational misrepair (strictly speaking, all three pathways involve recombinational events). The third pathway (Fig. 3C) represents the long-standing concepts of Revell’s exchange theory (reviews:[Savage, 1998; Edwards, 2002]). Here the initiating lesions are not outright radiogenic breaks that disrupt the continuity of chromosomes, and any breakage that does occur is a consequence of a subsequent enzymatic rejoining/misrejoining process.

Figure 3.
Standard models of exchange formation. The figure depicts radiation-induced DSBs (shown as gaps) and their misrejoining. Panel A describes a pathway where the two free ends of one break can either misrejoin independently of each other, at different genomic locations (Panel Aii), or act in concert with both free ends going to the same genomic location (Panel Ai). Due to the possibility of independent misrejoining, complex aberrations can arise readily and very complex aberrations can result (compare Fig. 2). Panel B indicates a different pathway. One essential difference is that a single radiation-induced DSB can lead to an aberration, perhaps by enzymatically-mediated homologous misrepair as shown. In the usual versions of this type of model, DSB free ends are constrained to act in concert during the recombinational event, as shown in panel B. This constraint leads to model predictions of a much smaller proportion of complex aberrations relative to simple ones than in the breakage-and-reunion case (see text). It also limits the type of aberrations that can arise. For example, in an mFISH experiment, any misrejoining must make either two color junctions between a particular pair of colors or none; consequently, the total number of color junctions between any given pair of colors should be an even number. Metaphases are observed where there are odd numbers of color junctions between some pairs of colors, an observation that favors pathway A over pathway B. Proponents of pathway B have, however, pointed out several confounding factors, including the following: (1) some rearranged chromatin pieces may be too short to observe, which would invalidate the even/odd argument; (2) the two pathways shown in A and B might be acting in parallel, so that some observed rearrangements result from one pathway and some from the other; (3) the constraint of two free ends acting in concert may be an incidental, not essential, feature of a 1-hit model. Because of such arguments, the question remains controversial, and quantitative model predictions of the ratio of complex to simple aberrations, which also bear on the differences between the pathways in A and in B, have drawn added attention. Panel C shows the Revell-type exchange-theory pathway. As in A, two radiation-induced lesions are required to initiate the exchange process. As in B, free ends of the same break are constrained to act in concert, restricting the type and frequency of complex aberrations.Chromosome localization and proximity effects
Regardless of the pathway, formation of chromosome exchanges involves having two or more different genomic loci in close proximity (Figs. 1-3). Consequently, chromosome geometry and large-scale chromatin architecture in the interphase nucleus influence aberration spectra. Conversely, aberration spectra can be used to probe chromosome localization and to help estimate the DSB interaction range, dependent on chromosome motion.[Brenner, 1990; Kellerer, 1978]
Some pioneering quantitative analyses of interphase chromosome geometry used radiation-induced chromosome aberrations (e.g.[Savage, 1973]), but until recently comparatively little direct information was available. The convoluted looping structure of interphase chromosomes defeated attempts to visualize chromatin architecture on the comparatively large scales (> 1 Mb) involved in aberration studies. Major progress made over the last decade has now shown that, at any one instant, any one chromosome is predominantly localized to a territory whose volume, in a human cell, is only a few percent of the volume of the whole nucleus (review:[Cremer, 2001])
The need for close spatial proximity of loci in an aberration-forming interaction, taken together with chromosome localization leads to a number of proximity effects, which can be quantified using computer modeling. Proximity effects include the following:
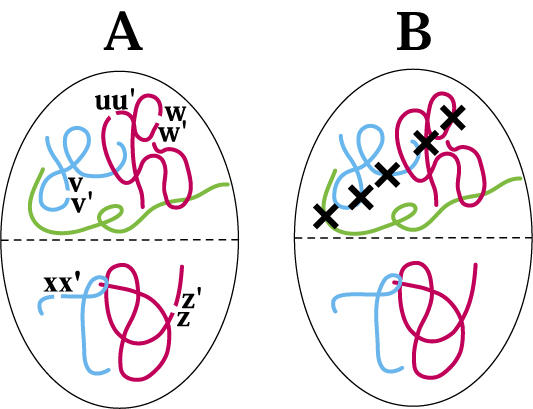
Figure 4.
Chromatin architecture, in relation to aberration spectra and radiation track structure. One way in which territory/proximity effects are seen is in a statistical bias for aberrations involving a single chromosome as compared to aberrations involving two different chromosomes. A. The basic phenomenon in simplified form. The panel schematically shows five G1 phase chromosomes in an interphase cell nucleus, localized to territories, with five DSBs. For sparsely ionizing radiation, such as gamma-rays, all five DSBs will usually (though not always) come from different radiation tracks, and usually be scattered at random throughout the nucleus.DSB free ends can interact only over a limited range, so proximity effects influence misrejoining. In the figure proximity effects can be visualized if we imagine that a free end can interact only with a free end in its own half of the nucleus, as indicated schematically by the dotted line. For example the free end u of DSB uu' can restitute with u' or misrejoin with v, v', w, or w', but not misrejoin with x, x', z, or z'.
In general, there will be a bias, relative to expectations based on randomness with all five DSBs capable of interacting, for forming a ring or inversion (Fig. 1B) compared to forming a translocation or a dicentric (Fig. 1A), i.e. a bias for one-chromosome aberrations as compared to two-chromosome aberrations. Thus in the diagram some reactions that can form dicentrics or translocations are allowed (e.g. the reaction xz, x'z', forming a translocation or dicentric, while the other three DSBs restitute as uu',vv', ww'; or the reaction vw, v'w', with restitutions uu', xx', zz'; etc.). But territory/proximity effects prohibit some dicentrics or translocations that could otherwise occur (e.g. ux, u'x'). In this diagram, territory/proximity effects have no effect on reactions capable of forming rings (i.e. u'w'). Overall, therefore, the ratio of dicentrics plus translocations to rings is smaller than it would be if territory/proximity effects were absent.
B. For the case of densely ionizing radiations, such as alpha particles, the five DSBs would typically be near the track of a single alpha particle, as indicated by the crosses, instead of being randomly located in the nucleus as in A. This difference in spatial DSB patterns leads to a different aberration spectrum, as described in the text.
Direct evidence indicates little overlap of chromosome territories.[Cremer, 2001] Correspondingly, recent results for the dependence of aberration frequency on chromosome DNA content support a picture where inter-chromosomal interactions occur mainly near the surface of territories.[Knehr, 1996; Cigarrán, 1998; Ostashevsky, 2000; Wu, 2001; Nakamura, 2001] However, the formation of multi-chromosome aberrations (e.g. Figs. 2 and 3Aii) suggests, to the contrary, considerable territorial overlap.[Sachs, 2000] One possible explanation for this discrepancy is preferential induction of aberrations in special locations outside the main territories, where loops from many different chromosomes may be close [Savage, 2000]. There is some evidence[Nelms, 1998] against an alternate explanation[Savage, 1998], of active transport of damaged chromatin to repair "factories" where DNA on different chromosomes is simultaneously processed. Motion observed on the scale of a whole chromosome territory (~1 μm) is constrained, rather slow, and apparently random[Edelmann, 2001; Kreth, 2001; Masuzawa, 2000; Marshall, 1997], though directed motion for a small portion of a chromosome is not decisively precluded.
Modeling aberration formation mechanisms quantitatively
In order to quantify proximity effects or other aspects of repair/misrepair mechanisms, and to make sense of the complicated data sets, computer modeling of aberration formation has become common.[Lucas, 1993; Sachs, 2000] Modern models of chromatin structure[Cremer, 2001; Münkel, 1999] can be integrated with simple models of DSB motion and misrejoining as well as with previously developed, sophisticated radiation track codes describing physical and radiochemical aspects of radiation.[Friedland, 1999; Chatterjee, 2001; Ottolenghi, 2001; Edwards, 2002]
Such aberration modeling is probabilistic, implemented by what are called Monte Carlo techniques, where the computer in effect "rolls dice" to give extremely detailed output. The time course of aberration formation is simulated in each cell. For example, suppose it is known that on average 1.5 DSBs are produced in any homologue of chromosome 1 in a given experiment. The computer first selects whether the number of DSBs for a copy of chromosome 1 in the first cell is 0, 1, 2, etc., either by using a random number generator together with an appropriate probability distribution or by using a probabilistic radiation-track code together with a polymer, random-walk geometric model of the chromosome. The specific location of the simulated DSB(s) on the chromosome is found. The other 45 chromosomes are then treated similarly, taking into account their geometry and DNA content. Repair/misrepair for all the DSBs is next simulated, as what is called a discrete-time Markov process, by an algorithm probabilistically favoring misrejoining of those DSB free ends that are spatially close to each other. The result is a simulated configuration of rearranged chromosomes in the first cell. Simulating the scoring system (such as mFISH) used in the experiment then gives the observable aberration pattern for that simulated cell; a simple example might be that the cell contains just one dicentric, involving chromosomes 4 and X. Iterating, thousands or millions of individual cells are simulated one by one, each with its own aberration pattern. The results are then compared to experimentally observed aberration spectra and to dose-response relationships for aberration frequencies.
This quantitative, probabilistic approach requires making explicit the basic assumptions (e.g. which aberration formation pathway is being considered and what geometric model is being used for chromosomes). It systematically emphasizes dominant processes and likely outcomes, appropriately discounting, without completely ignoring, minor contributory pathways and a large number of possible but highly unlikely aberration patterns. The approach uses randomness assumptions[Savage, 1982] on DSB distribution in the genome and on misrejoining to test mechanistic models quantitatively, using a minimum number of adjustable parameters. Randomness holds to reasonable approximation (reviews:[Johnson, 1999; Sachs, 2000]), though there are some reports of "hot spots" or other deviations from randomness (e.g.[Day, 1998; Kilburn, 2001; Knehr, 1996; Cigarrán, 1998]).
Based on such biophysical modeling, on analyzing aberration dose-response (discussed below), and on evidence concerning likely molecular mechanisms, we believe that breakage-and-reunion is the dominant pathway for aberrations produced in mammalian cells following exposure to ionizing radiation during G0/G1.[Cornforth, 2001; Sachs, 2000; Cornforth, 1998; Hoeijmakers, 2001; Richardson, 2000] One main reason is that in the simulations the other mechanisms described in Fig. 3 are unable to reproduce the full richness in aberration spectra that are observed experimentally, especially the frequency and extent of complex aberrations. However, the modeling does not preclude some admixture of the other pathways, and the situation remains controversial.[Cucinotta, 2000; Edwards, 2002]
An excess of very complex aberrations?
If one accepts the breakage-and-reunion model and assumes that all reactions are complete (i.e. that no DSB free ends are left over at the end), one can prove that any aberration-producing reaction can be uniquely decomposed into irreducible reactions called cycles.([Bafna, 1996; Sachs, 1999; Cornforth, 2001]) The order of a cycle quantifies aberration complexity. For example Figs. 1Ai-iii, 1Bi, 1Bii, and 3Ai all correspond to cycles of order 2; Fig. 1Aiii is the result of two cycles of order 1 (restitutions); Fig 3Aii shows a cycle of order 3; and analysis of Fig. 2 indicates that one of the reactions was a cycle of order 6. mFISH data shows cycles of unexpectedly high order and cells with unusually large numbers of different chromosomes taking part in aberrations.[Greulich, 2000; Loucas, 2001] Biophysical modeling suggests that an additional mechanism may be operating, to generate the most complex rearrangements seen.[Sachs, 2000; Vázquez, 2002] This could perhaps be an early onset of chromsomal instability.
From mechanisms to predicted dose and dose-rate dependencies
Apart from insights into nuclear architecture and into DNA repair/misrepair, mechanistic understanding of chromosome aberration formation is useful in various applications of radiobiology. Almost always, the key questions in applications are how radiation effects depend on dose and on dose rate. Understanding aberration formation mechanisms helps answer these two questions. Experiments, and models using ordinary differential equations and/or stochastic-process theory, have over the years uncovered some rather general rules about dose and dose-rate dependence for different mechanisms (reviews:[Sachs, 1997; Stewart, 2001]). The rules are based primarily on a distinction between "1-track action" and "2-track action" (Fig. 5), where 2-track action involves the interaction of uncorrelated damage from 2 different primary radiation tracks (such as two different x-ray photons), so that its dose and dose-rate dependencies are more complicated than for 1-track (i.e. intra-track) action. Despite some exceptions, and the fact that they are approximations rather than exact statements, the rules cover a large variety of cases. They often hold to good approximation, not only for simple chromosome aberrations, as illustrated in Fig. 5, but also for many other kinds of ionizing radiation damage. The rules are the following.
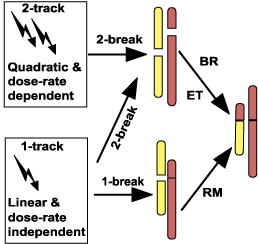
Figure 5. Dose and dose-rate dependence of aberration formation. The figure shows different pathways for producing the simple chromosome aberration on the right. It summarizes basic rules for the way in which the yield of simple aberrations depends on dose and dose rate.
Applying these rules to the chromosome aberration formation pathways discussed earlier, one sees from Fig. 5 that the breakage-and-reunion (BR) and the Revell-type exchange theory (ET) mechanisms, both of which require two different radiation-induced breaks to initiate an exchange (Fig. 3), lead to a mixture of linear and quadratic dose-dependence. The linear component occurs because a single track sometimes produces two or more DSBs (Fig. 4). In this linear-quadratic response the linear term dominates at low doses, and the quadratic term dominates at higher doses.[Edwards, 2002] One also sees from the figure that the recombinational misrepair (RM) mechanism (Fig. 3) usually leads to a linear dose-dependence that is independent of dose rate.
For brevity, some complications are omitted from Fig. 5, including the following.
Medical and public health aspects
Despite these complications, the rules of Fig. 5 have important applications to biodosimetry, risk estimation, and radiotherapy treatment planning.
In applications to biodosimetry the goal is usually to infer dose retrospectively from the level of chromosome aberrations in an individual's peripheral blood lymphocytes. This procedure requires knowing how aberration frequencies vary with dose for the particular radiation type and exposure conditions involved. Under the breakage-and-reunion scenario described above, radiation induced dicentric or translocation frequency is expected to have, for sparsely ionizing radiation in the relevant dose-range, a linear-quadratic dependence on dose (Fig. 5). In vitro, this expected linear-quadratic dependence is in fact observed.[Edwards, 2002; Bauchinger, 1998] Densely ionizing radiation, on the other hand, operates almost exclusively via intra-track action (Fig. 4) over the relevant dose range, so near-linearity in dose is expected (Fig. 5), and is observed.[Edwards, 2002; Bauchinger, 1998]
In addition, because of the interplay between chromatin geometry and radiation track structure (Fig. 4), the spectrum of aberration types is expected to be different for densely ionizing radiation.[Brenner, 1994] A different spectrum is indeed observed in vitro: there are higher frequencies of aberrations involving several exchange breakpoints within the same chromosome arm, compared to inter-chromosomal interactions; and at low doses there is a higher frequency of complex aberrations compared to simple ones.[Bauchinger, 1998; Boei, 2001; Sabatier, 1987; Anderson, 2000] Because of such tell-tale differences, retrospective biodosimetery should eventually be able to identify the type of radiation as well as the dose received.
Another potential application involves estimating cancer risks from radiation exposure. A long standing problem has been that the doses of primary interest are too small to produce quantifiable ¾ or, often, even detectable ¾ biological effects, either experimentally or even epidemiologically. And yet, there are serious concerns over the effects of such low doses when acting on very large populations. Major uncertainties in low-dose and low dose-rate experimental estimates, together with the major health and economic issues involved, have made this area highly contentious. Biophysical, mechanistic models of radiation damage, such as the models outlined above, though themselves controversial, are one of the few hopes for extrapolating measurable risks appropriately to lower doses. This approach to risk estimation is valid only to the extent that dose and dose-rate dependence of radiation carcinogenesis parallel those of aberration formation. Currently risk estimate extrapolations from higher doses, with consideration given to effects of dose-rate, are based on the linear-quadratic model (Fig. 5), in part motivated by results on chromosome aberrations.[Fry, 1998; NCRP, 2001] There is also good evidence for a causal link between translocations and certain cancers, especially leukemias; on the other hand, the radiogenesis of some solid tumors may be more closely related to other forms of radiation damage, having different dose- and dose-rate dependency.[Little, 2000]
A third application concerns treatment of tumors with radiation. Much of the tumor cell killing, and the undesired side effect of killing surrounding normal cells, probably comes from production of dicentrics and centric rings (Fig. 1).[Bedford, 1991; Franken, 1999] A linear-quadratic dose-response together with a dose-rate effect, suggested by chromosome aberration results and modeling (Fig. 5), is confirmed by more direct clinical data and forms the basis of modern, biologically based treatment planning for tumor radiotherapy.[Baumann, 2001; Hall, 2000] In addition, aberration-based predictive assays for sensitivity of normal tissues surrounding a tumor offer promise of individualized treatment.[Coco Martin, 1999; Barber, 2000]
Conclusions and prospects
Recent technological advances such as mFISH and spectral karyotyping have led to an explosive increase in cytogenetic data, which, together with computer-assisted modeling, allow new insights into the formation of radiation-induced chromosome aberrations. For mammalian cells, the weight of the evidence¾ on complexity of aberrations, on underlying molecular mechanisms, and on dose-response/dose-rate relationships ¾ favors a breakage-and-reunion mechanism during G0/G1, involving non-homologous end-joining. The chromosome geometry picture emerging, from radiobiological and other data, is one of localized chromosomes, overlapping and interacting with each other mainly at territory surfaces or via loops protruding far from the home territories. For the most part whole-chromosome territories seem to be almost randomly located with respect to each other. The randomness is modulated by some more systematic associations, but these are typically weak, transient, variable from cell to cell, or highly localized in the genome rather than being firm whole-chromosome associations, so they show up in aberration experiments as a statistical bias for extra exchanges, set against a backdrop where any chromosome can undergo exchanges with any other.
Better quantitative characterizations of DNA repair/misrepair mechanisms and of chromosome geometry will surely emerge from currently ongoing aberration work, but some other areas call for new initiatives. One issue that has not to date received as much attention as it deserves is the interrelation between aberrations analyzed cytogenetically and mutations identified by using selection for specific missing gene products. There is a considerable consensus that most "large" mutations (e.g. total deletions of the HPRT gene) are formed by essentially the same pairwise misrepair mechanism(s) as intrachromosomal exchange aberrations (review:[Costes, 2001]), but more systematic attempts to interrelate aberrations and mutations quantitatively are needed. As well, more experimental evidence and quantitative modeling on the relation between gene expression microarray data and aberration data in radiobiology is needed. Applications of aberrations to retrospective biodosimetry and tumor radiotherapy will no doubt benefit from continued technical improvements in cytogentic techniques, but for applications to radiation risk estimation technical improvements by themselves do not hold out as much promise. Probably nothing less than a conceptual breakthrough as regards radiation carcinogenesis can lead to credible low-dose risk estimates.
Acknowledgements
We are grateful to D.J. Brenner and P. Hahnfeldt for wide-ranging discussions.
References
Anderson G, D Stoler and B Brenner (2001), Cancer: the evolved consequence of a destabilized genome, Bioessays 23, 1037-1046.
Anderson RM, SJ Marsden, EG Wright, MA Kadhim, DT Goodhead and CS Griffin (2000), Complex chromosome aberrations in peripheral blood lymphocytes as a potential biomarker of exposure to high-LET alpha-particles, International Journal of Radiation Biology. 76, 31-42.
Bafna V and PA Pevzner (1996), Genome Rearrangements and Sorting by Reversals, Siam Journal on Computing 25, 272-289.
Barber JBP, W Burrill, AR Spreadborough, E Levine, C Warren, AE Kiltie, SA Roberts and D Scott (2000), Relationship between in vitro chromosomal radiosensitivity of peripheral blood lymphocytes and the expression of normal tissue damage following radiotherapy for breast cancer, Radiotherapy & Oncology 55, 179-186.
Bauchinger M (1998), Retrospective dose reconstruction of human radiation exposure by FISH/chromosome painting, Mutation Research 404, 89-96.
Bauchinger M and E Schmid (1998), LET dependence of yield ratios of radiation-induced intra- and interchromosomal aberrations in human lymphocytes, International Journal of Radiation Biology 74, 17-25.
Baumann M, SM Bentzen, W Doerr, MC Joiner, M Saunders, IF Tannock and HD Thames (2001), The translational research chain: Is it delivering the goods?, International Journal of Radiation Oncology Biology Physics 49, 345-351.
Bedford JS (1991), Sublethal damage, potentially lethal damage, and chromosomal aberrations in mammalian cells exposed to ionizing radiations, International Journal of Radiation Oncology Biology Physics 21, 1457-1469.
Bode J, C Benham, E Ernst, A Knopp, R Marschalek, R Strick and P Strissel (2000), Fatal connections: When DNA ends meet on the nuclear matrix, Journal of Cellular Biochemistry Supplement 35, 3-22.
Boei JJWA, S Vermeulen and AT Natarajan (2000), Analysis of radiation-induced chromosomal aberrations using telomeric and centromeric PNA probes, International Journal of Radiation Biology 76, 163-167.
Boei JJWA, S Vermeulen, LHF Mullenders and AT Natarajan (2001), Impact of radiation quality on the spectrum of induced chromosome exchange aberrations, International Journal of Radiation Biology 77, 847-857.
Boyle S, S Gilchrist, JM Bridger, NL Mahy, JA Ellis and WA Bickmore (2001), The spatial organization of human chromosomes within the nuclei of normal and emerin-mutant cells, Human Molecular Genetics 10, 211-219.
Brenner D (1990), Track structure, lesion development, and cell survival, Radiation Research 124, S29-37.
Brenner D, J Ward and R Sachs (1994), Track structure, chromosome geometry and chromosome aberrations, Basic Life Sciences 63, 93-113.
Cafourkova A, E Lukasova, S Kozubek, M Kozubek, RD Govorun, I Koutna, E Bartova, M Skalnikova, P Jirsova, R Pasekova and EA Krasavin (2001), Exchange aberrations among 11 chromosomes of human lymphocytes induced by gamma-rays, International Journal of Radiation Biology 77, 419-429.
Chatterjee A, W Holley, B Rydberg, S Mian and RD Alexander (2001), Theoretical modeling of DNA damages and cellular responses, Physica Medica XVII, Supplement 1, 59-61.
Cigarrán S, L Barrios, JF Barquinero, MR Caballín, M Ribas and J Egozcue (1998), Relationship between the DNA content of human chromosomes and their involvement in radiation-induced structural aberrations, analysed by painting, International Journal of Radiation Biology 74, 449-55.
Coco Martin JM, E Mooren, C Ottenheim, W Burrill, MI Nunez, D Sprong, H Bartelink and AC Begg (1999), Potential of radiation-induced chromosome aberrations to predict radiosensitivity in human tumour cells, International Journal of Radiation Biology 75, 1161-1168.
Cornforth MN (1998). "Radiation-induced damage and the formation of chromosomal aberrations", Humana Press, Totowa, N.J.
Cornforth MN (2001), Analyzing radiation-induced complex chromosome rearrangements by combinatorial painting, Radiation Research 155, 643-659.
Costes S, R Sachs, L Hlatky, D Vannais, C Waldren and B Fouladi (2001), Low-LET, large-mutation spectra at hemizygous loci: evidence for intrachromosomal proximity effects, Radiation Research 156, 545-557.
Cremer M, J von Hase, T Volm, A Brero, G Kreth, J Walter, C Fischer, I Solovei, C Cremer and T Cremer (2001), Non-random radial higher-order chromatin arrangements in nuclei of diploid human cells, Chromosome Research 9, 541-567.
Cremer T and C Cremer (2001), Chromosome territories, nuclear architecture and gene regulation in mammalian cells, Nature Review Genetics 2, 292-301.
Cucinotta FA, H Nikjoo, P O'Neill and DT Goodhead (2000), Kinetics of DSB rejoining and formation of simple chromosome exchange aberrations., International Journal of Radiation Biology 76, 1463-1474.
Day JP, CL Limoli and WF Morgan (1998), Recombination involving interstitial telomere repeat-like sequences promotes chromosomal instability in Chinese hamster cells, Carcinogenesis (Oxford) 19, 259-266.
Edelmann P, H Bornfleth, D Zink, T Cremer and C Cremer (2001), Morphology and dynamics of chromosome territories in living cells, Biochimica et Biophysica Acta 1551, M29-M40.
Edwards AA (1997), The use of chromosomal aberrations in human lymphocytes for biological dosimetry, Radiation Research 148, S39-S44.
Edwards AA (2002), Modeling radiation-induced chromosome aberrations, International Journal of Radiation Biology 78, 000-000.
Fauth C and MR Speicher (2001), Classifying by colors: FISH-based genome analysis, Cytogenetics and Cell Genetics 93, 1-10.
Franken NAP, P Ruurs, G Ludwikow, C Van Bree, JBA Kipp, F Darroudi and GW Barendsen (1999), Correlation between cell reproductive death and chromosome aberrations assessed by FISH for low and high doses of radiation and sensitization by iodo-deoxyuridine in human SW-1573 cells, International Journal of Radiation Biology 75, 293-299.
Friedland W, P Jacob, HG Paretzke, M Merzagora and A Ottolenghi (1999), Simulation of DNA fragment distributions after irradiation with photons, Radiation and Environmental Biophysics 38, 39-47.
Fry RJ, A Grosovsky, PC Hanawalt, RF Jostes, JB Little, WF Morgan, NL Oleinick and RL Ullrich (1998), The Impact of Biology on Risk Assessment--workshop of the National Research Council's Board on Radiation Effects Research. July 21-22, 1997, National Academy of Sciences, Washington, DC, Radiation Research 150, 695-705.
Gisselsson D (2001). Atlas of Genetics and Cytogenetics in Oncology and Haematology.
Goodhead DT, J Thacker and R Cox (1993), Weiss Lecture. Effects of radiations of different qualities on cells: molecular mechanisms of damage and repair, International Journal of Radiation Biology 63, 543-56.
Greulich KM, L Kreja, B Heinze, AP Rhein, H-UG Weier, M Brückner, P Fuchs and M Molls (2000), Rapid detection of radiation-induced chromosomal aberrations in lymphocytes and hematopoietic progenitor cells by mFISH, Mutation Research 452, 73-81.
Grosovsky AJ, KK Parks, CR Giver and SL Nelson (1996), Clonal analysis of delayed karyotypic abnormalities and gene mutations in radiation-induced genetic instability, Molecular and Cellular Biology 16, 6252-62.
Hall EJ (2000). "Radiobiology for the radiologist" 5 ed., Lippincott Williams & Wilkins., Philadelphia.
Hoeijmakers JHJ (2001), Genome maintenance mechanisms for preventing cancer, Nature 411, 366-374.
Johnson KL, DJ Brenner, J Nath, JD Tucker and CR Geard (1999), Radiation-induced breakpoint misrejoining in human chromosomes: random or non-random?, International Journal of Radiation Biology 75, 131-41.
Karhu R, M Ahlstedt-Soini, M Bittner, P Meltzer, JM Trent and JJ Isola (2001), Chromosome arm-specific multicolor FISH, Genes Chromosomes & Cancer 30, 105-109.
Kellerer A and H Rossi (1978), A generalized formulation of dual radiation action., Radiation Research 75, 471-488.
Kilburn AE, MJ Shea, RG Sargent and JH Wilson (2001), Insertion of a telomere repeat sequence into a mammalian gene causes chromosome instability, Molecular and Cellular Biology 21, 126-135.
Knehr S, H Zitzelsberger, H Braselmann, U Nahrstedt and M Bauchinger (1996), Chromosome analysis by fluorescence in situ hybridization: further indications for a non-DNA-proportional involvement of single chromosomes in radiation-induced structural aberrations, International Journal of Radiation Biology 70, 385-392.
Kreth G, P Edelmann, T Cremer and C Cremer (2001), Towards the computer modelling of dynamic nuclear organization, Analytical Cellular Pathology 22, 40.
Lee C, D Gisselsson, C Jin, A Nordgren, DO Ferguson, E Blennow, JA Fletcher and CC Morton (2001), Limitations of chromosome classification by multicolor karyotyping, American Journal of Human Genetics 68, 1043-1047.
Little MP (2000), A comparison of the degree of curvature in the cancer incidence dose-response in Japanese atomic bomb survivors with that in chromosome aberrations measured in vitro, International Journal of Radiation Biology 76, 1365-1375.
Loucas BD and MN Cornforth (2001), Complex chromosome exchanges induced by gamma rays in human lymphocytes: an mFISH study, Radiation Research 155, 660-671.
Lucas JN and RK Sachs (1993), Using three-color chromosome painting to test chromosome aberration models, Proceedings of the National Academy of Sciences of the United States of America 90, 1484-1487.
Marshall WF, A Straight, JF Marko, J Swedlow, A Dernburg, A Belmont, AW Murray, DA Agard and JW Sedat (1997), Interphase chromosomes undergo constrained diffusional motion in living cells, Current Biology 7, 930-939.
Masuzawa N, Y Urata, K Yagi and T Ashihara (2000), Constrained, random, and independent motion of texas-red-labeled chromatin in living interphase PtK2 cells, Acta Histochemica Et Cytochemica 33, 419-427.
Mitelman F, B Johansson and F Mertens (2002).
Münkel C, R Eils, S Dietzel, D Zink, C Mehring, G Wedeman, T Cremer and J Langowski (1999), Compartmentalization of interphase chromosomes observed in simulation and experiment, Journal of Molecular Biology 285, 1053-1065.
Nakamura N, K Ohtaki, Y Kodama, M Nakano, M Itoh, AA Awa and JB Cologne (2001), Randomness of Radiation-induced Translocations Depends on Chromosome Length or Surface Area as Target, REREF Update 12, 12-15.
Nakano M, Y Kodama, K Ohtaki, M Itoh, R Delongchamp, AA Awa and N Nakamura (2001), Detection of stable chromosome aberrations by FISH in A-bomb survivors: Comparison with previous solid Giemsa staining data on the same 230 individuals, International Journal of Radiation Biology 77, 971-977.
NCRP (2001). "Evaluation of the linear nonthreshold dose-response model for ionizing radiation", NCRP, Bethesda.
Nelms BE, RS Maser, JF Mackay, MG Lagally and JHJ Petrini (1998), In situ visualization of DNA double-strand break repair in human fibroblasts, Science (Washington D C) 280, 590-592.
Nikiforova MN, JR Stringer, R Blough, M Medvedovic, JA Fagin and YE Nikiforov (2000), Proximity of chromosomal loci that participate in radiation-induced rearrangements in human cells, Science 290, 138-141.
Ostashevsky JY (2000), Higher-order structure of interphase chromosomes and radiation-induced chromosomal exchange aberrations, International Journal of Radiation Biology 76, 1179-1187.
Ottolenghi A, F Ballarini and M Biaggi (2001), Modelling chromosomal aberration induction by ionising radiation: the influence of interphase chromosome architecture, Advances in Space Research 27, 369-82.
Padilla-Nash HM, K Heselmeyer-Haddad, D Wangsa, H Zhang, BM Ghadimi, M Macville, M Augustus, E Schrock, E Hilgenfeld and T Ried (2001), Jumping translocations are common in solid tumor cell lines and result in recurrent fusions of whole chromosome arms, Genes Chromosomes & Cancer 30, 349-363.
Pfeiffer P, W Goedecke and G Obe (2000), Mechanisms of DNA double-strand break repair and their potential to induce chromosomal aberrations, Mutagenesis 15, 289-302.
Pinkel D, J Gray, B Trask, G van den Engh, J Fuscoe and H van Dekken (1986), Cytogenetic analysis by in situ hybridization with fluorescently labeled nucleic acid probes., Cold Spring Harbor Symposia on Quantitative Biology 51, 151-7.
Radivoyevitch T, DG Hoel, AM Chen and RK Sachs (1998), Misrejoining of double-strand breaks after X irradiation: relating moderate to very high doses by a Markov model, Radiation Research 149, 59-67.
Radivoyevitch T, S Kozubek and R Sachs (2001), Biologically based risk estimation for radiation-induced CML - Inferences from BCR and ABL geometric distributions., Radiation and Environmental Biophysics 40, 1-9.
Richardson C and M Jasin (2000), Frequent chromosomal translocations induced by DNA double-strand breaks, Nature 405, 697-700.
Rossi H and M Zaider (1996). "Microdosimetry and its applications", Springer, New York.
Rothkamm K, M Kuehne, PA Jeggo and M Loebrich (2001), Radiation-induced genomic rearrangements formed by nonhomologous end-joining of DNA double-strand breaks, Cancer Research 61, 3886-3893.
Sabatier L, W Al Achkar, F Hoffschir, C Luccioni and B Dutrillaux (1987), Qualitative study of chromosomal lesions induced by neutrons and neon ions in human lymphocytes at G0 phase, Mutat Res 178, 91-7.
Sachs RK, DJ Brenner, AM Chen, P Hahnfeldt and LR Hlatky (1997a), Intra-arm and interarm chromosome intrachanges: tools for probing the geometry and dynamics of chromatin, Radiation Research 148, 330-40.
Sachs RK, P Hahnfeld and DJ Brenner (1997b), The link between low-LET dose-response relations and the underlying kinetics of damage production/repair/misrepair, International Journal of Radiation Biology 72, 351-74.
Sachs RK, AM Chen, PJ Simpson, LR Hlatky, P Hahnfeldt and JR Savage (1999), Clustering of radiation-produced breaks along chromosomes: modelling the effects on chromosome aberrations, International Journal of Radiation Biology 75, 657-72.
Sachs RK, D Levy, AM Chen, PJ Simpson, MN Cornforth, EA Ingerman, P Hahnfeldt and LR Hlatky (2000a), Random breakage and reunion chromosome aberration formation model; an interaction-distance version based on chromatin geometry, International Journal of Radiation Biology 76, 1579-88.
Sachs RK, A Rogoff, AM Chen, PJ Simpson, JR Savage, P Hahnfeldt and LR Hlatky (2000b), Underprediction of visibly complex chromosome aberrations by a recombinational-repair ('one-hit') model, International Journal of Radiation Biology 76, 129-48.
Saracoglu K, J Brown, L Kearney, S Uhrig, J Azofeifa, C Fauth, MR Speicher and R Eils (2001), New concepts to improve resolution and sensitivity of molecular cytogenetic diagnostics by multicolor fluorescence in situ hybridization, Cytometry 44, 7-15.
Savage J and D Papworth (1973), The relationship of radiation-induced dicentric yield to chromosome arm number., Mutation Research 19, 139-43.
Savage JRK and DG Papworth (1982), Frequency and distribution studies of asymmetrical versus symmetrical chromosome aberrations, Mutation Research 95, 7-18.
Savage JRK (1998), A brief survey of aberration origin theories, Mutation Research 404, 139-147.
Savage JRK (2000), Enhanced Perspective: Proximity Matters, Science 290, 62-63.
Schrock E and H Padilla-Nash (2000), Spectral karyotyping and multicolor fluorescence in situ hybridization reveal new tumor-specific chromosomal aberrations., Seminars in Hematology 37, 334-47.
Speicher MR, SG Ballard and DC Ward (1996), Karyotyping human chromosomes by combinatorial multi-fluor FISH, Nature Genetics 12, 368-375.
Stewart RD (2001), Two-lesion kinetic model of double-strand break rejoining and cell killing, Radiation Research 156, 365-378.
Thacker J (1999), Repair of ionizing radiation damage in mammalian cells. Alternative pathways and their fidelity, Comptes rendus de l'academie des sciences. serie iii, sciences de la vie 322, 103-8.
Uhrig S, S Schuffenhauer, C Fauth, A Wirtz, C Daumer-Haas, C Apacik, M Cohen, J Mueller-Navia, T Cremer, J Murken and MR Speicher (1999), Multiplex-FISH for pre- and postnatal diagnostic applications, American Journal of Human Genetics 65, 448-462.
Vázquez M, K Greulich-Bode, J Arsuaga, M Cornforth, M Brückner, R Sachs, P Hahnfeldt, M Molls and L Hlatky (2002), Computer analysis of mFISH chromosome aberration data uncovers an excess of very complex metaphases. Accepted for publication., International Journal of Radiation Biology 000, 000-000.
Watson GE, SA Lorimore, DA Macdonald and EG Wright (2000), Chromosomal instability in unirradiated cells induced in vivo by a bystander effect of ionizing radiation, Cancer Research 60, 5608-5611.
Wiegant J, V Bezrookove, C Rosenberg, HJ Tanke, AK Raap, H Zhang, M Bittner, JM Trent and P Meltzer (2000), Differentially painting human chromosome arms with combined binary ratio-labeling fluorescence in situ hybridization, Genome Research 10, 861-865.
Wu H, M Durante and JN Lucas (2001), Relationship between radiation-induced aberrations in individual chromosomes and their DNA content: Effects of interaction distance, International Journal of Radiation Biology 77, 781-786.